
فتوسنتز
فتوسنتز فرایندی است که گیاهان و سایرارگانیسمها از آن برای تبدیل انرژی نوری به انرژی شیمیایی استفاده میکنند. این انرژی شیمیایی در کربوهیدراتهایی ذخیره میشود که در مواقع فعالیتهای متابولیسمی از این قندها، طی فرایند تنفس سلولی برای تامین انرژی استفاده میشوند.

فتوسنتز، در بیشتر گیاهان، جلبکها و باکتریهای فتوسنتز کننده انجام میشود که به این ارگانیسم فتواتوتروف میگویند؛ به این معنا که این ارگانیسمها میتوانند با انرژیای که از نور میگیرند، منبع غذاییشان را تامین کنند. فتوسنتز تا حد زیادی مسئول تولید و حفظ اکسیژن جو زمین است و بیشتر انرژی مورد نیاز برای زندگی در زمین را تامین میکند. با این وجود، فتوسنتز درهر گونهای نسبت به گونه دیگر به طور متفاوت اتفاق میافتد.
فرایند فتوسنتز زمانی آغاز میشود که انرژی نوری توسط پروتئینهایی به نام مرکز واکنش، که حاوی رنگیزههای سبز کلروفیل است جذب شود، در گیاهان این پروتئینها درون اندامکهایی به نام کلروپلاست ذخیره میشوند که در سلولهای برگ به فراوانی یافت میشود در حالی که در باکتریها این پروتئینها درغشای پلاسمایی جا داده شدهاند.
انواع فتوسنتز در گیاهان
به طور کلی دو نوع فرایند فتوسنتز وجود دارد:
- فتوسنتز اکسیژنزا
- غیر اکسیژنزا
با اینکه اصول کلی فتوسنتز غیر اکسیژنزا و اکسیژنزا بسیار شبیه به هم است؛ اما فتوسنتز اکسیژنزا رایجتر میباشد و در گیاهان، جلبکها و سیانوباکتریها دیده میشود. در طول فتوسنتز اکسیژن زا، انرژی نور، الکترونها را از آب به کربن دی اکسید منتقل میکند تا کربوهیدرات تولید شود به این صورت که کربن دی اکسید با دریافت الکترون کاهش یافته و آب با از دست دادن الکترون اکسید میشود در نهایت اکسیژن به همراه کربوهیدرات تولید خواهد شد.
فتوسنتز اکسیژنزا با دریافت کربن دی اکسید تولید شده توسط ارگانیسمهای تنفسی و وارد کردن مجدد اکسیژن به اتمسفر، به عنوان متعادل کننده تنفس عمل میکند. از سوی دیگر، در فتوسنتز غیر اکسیژنزا، که به طور معمول در باکتریهایی مثل باکتریهای ارغوانی و باکتریهای سبز گوگردی اتفاق میافتد، از الکترون دهندههایی غیر از آب استفاده میشود؛ مثلاً در باکتریهای گوگردی سبز، عمدتاً از هیدروژن سولفید به عنوان الکترون دهنده استفاده میشود.
هر دو نوع فتوسنتز پیچیده و چند مرحلهای هستند اما روند کلی فتوسنتز اکسیژنزا را میتوان در معادله زیر خلاصه کرد:
۶CO۲ + ۱۲H۲O + نوری انرژی → C۶H۱۲O۶ + ۶CO۲ + ۶H۲O
فرآیند فتوسنتز
واکنشهای فتوسنتز در گیاهان به دو دسته وابسته به نور و مستقل از نور تقسیم میشود که هر دو این واکنشها در کلروپلاست رخ میدهد. کلروپلاست اندامکی درون سلولهای گیاهان و برخی جلبکها است، که فتوسنتز در آن انجام میشود، این اندامک دارای دو غشای بیرونی و درونی است که از هم فاصله دارند و بخش درونی آن استروما نام دارد؛ تیلاکوئیدها ساختارهایی کیسهای شکل از جنس غشای سلولاند که درون استروما قرار گرفته اند.
واکنشهای وابسته به نور درون تیلاکوئیدها و واکنشهای مستقل از نور در استروما رخ میدهد. واکنشهای وابسته به نور (واکنشهای نوری): هنگامی که فوتونهای نور به مرکز واکنش برخورد میکنند، کلروفیل الکترون آزاد میکند، الکترون آزاد شده با عبور از یک زنجیره انتقال الکترون انرژی مورد نیاز برای تهیه ATP و NADH را تامین میکند. کلروفیلی که الکترون از دست داده است، با گرفتن الکترون از آب و آزاد کردن اکسیژن، کمبود الکترون خود را جبران میکند.
واکنشهای مستقل از نور (واکنشهای تاریکی، چرخه کالوین): واکنشهای نوری، ATP و NADH را تولید میکنند که منابع غنی انرژی هستند، انجام واکنشهای تاریکی، وابسته به ATP و NADH حاصل از واکنشهای نوری است. اتمهای کربن موجود در دی اکسید کربن، با قرار گرفتن در چرخه کالوین به شکل قندهای سه کربنی تثبیت میشوند و در نهایت بخشی از این قندها برای تولید گلوکز و از بخش دیگر آن برای شروع مجدد چرخه کالوین استفاده میشود.
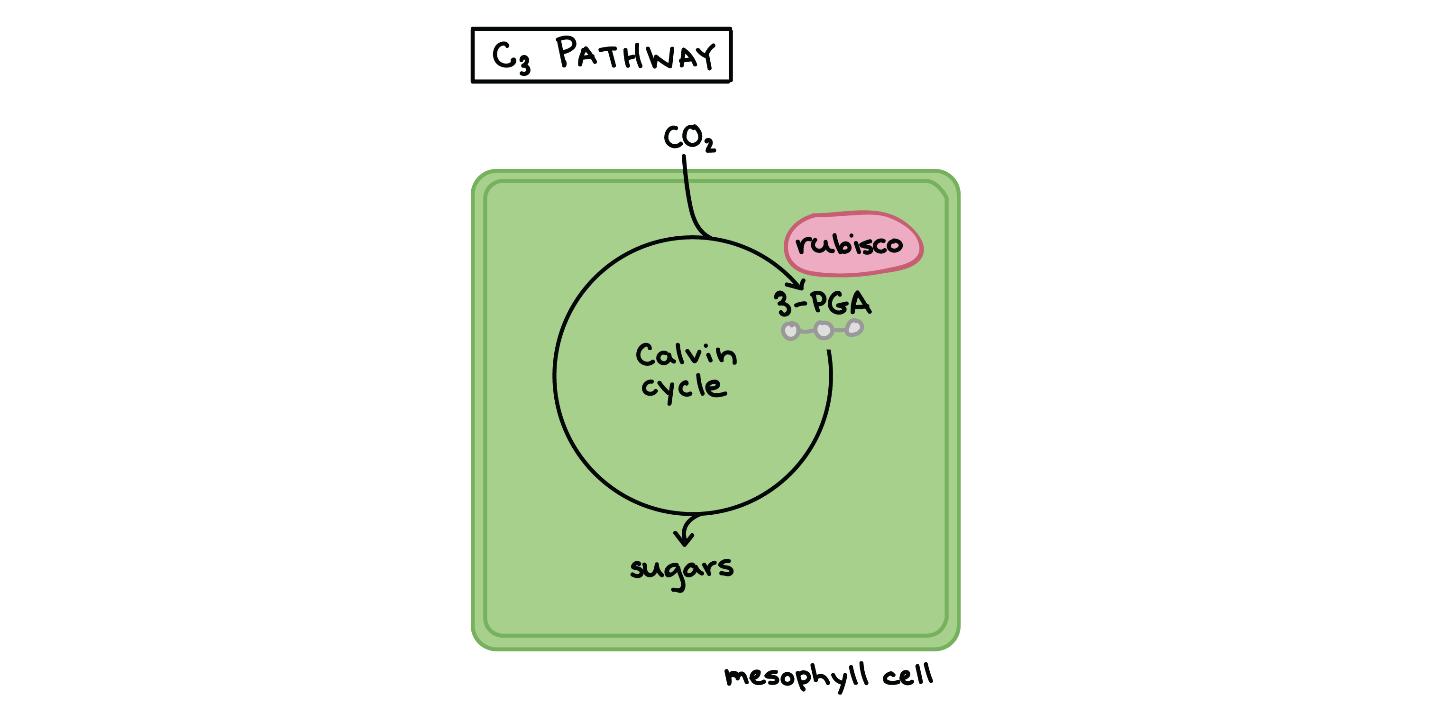
اولین مرحله از چرخه کالوین، تثبیت کربن دی اکسید توسط آنزیم روبیسکو است، روبیسکو آنزیمی است که هم به عنوان کربوکسیلاز و هم به عنوان اکسیژناز فعالیت میکند. نقش کربوکسیلازی یا اکسیژنازی این آنزیم به میزان کربن دی اکسید و اکسیژن موجود در محیط عملکرد آن بستگی دارد. گیاهانی که فقط از این مکانیزم استاندارد برای تثبیت کربن استفاده میکنند، C۳ نام دارند. این واکنش برای تهیه ترکیب ۳-PGA به کار میرود.
در مرحله دوم، ATP و NADPH با استفاده از انرژی ذخیره شده خود، مولکولهای ADP و +NAD را بازتولید وترکیب سه کربنی ۳-PGA را به یک ترکیب سه کربنی دیگر به نام G3P تبدیل میکنند. مولکولهای ADP و +NAD، ناشی از این واکنش کاهشی، به واکنشهای وابسته به نور برمی گردند تا دوباره انرژی بگیرند.
در آخرین مرحله، یکی از مولکولهای G3P چرخه کالوین را ترک میکند تا در تشکیل مولکول کربوهیدرات، که معمولاً گلوکز است ( C۶H۱۲O۶ )، کمک کند. از آنجا که مولکول کربوهیدرات شش اتم کربن دارد، برای ساختن یک مولکول کربوهیدرات شش دور از چرخه کالوین طول میکشد. مولکولهای G3P باقیمانده، RuBP را بازسازی میکنند، تا سیستم را قادر سازد که برای مرحله تثبیت کربن آماده شود. ATP همچنین در بازسازی RuBP استفاده میشود. فتوسنتز از نوع C۳ در۸۵٪ گیاهان انجام میشود.
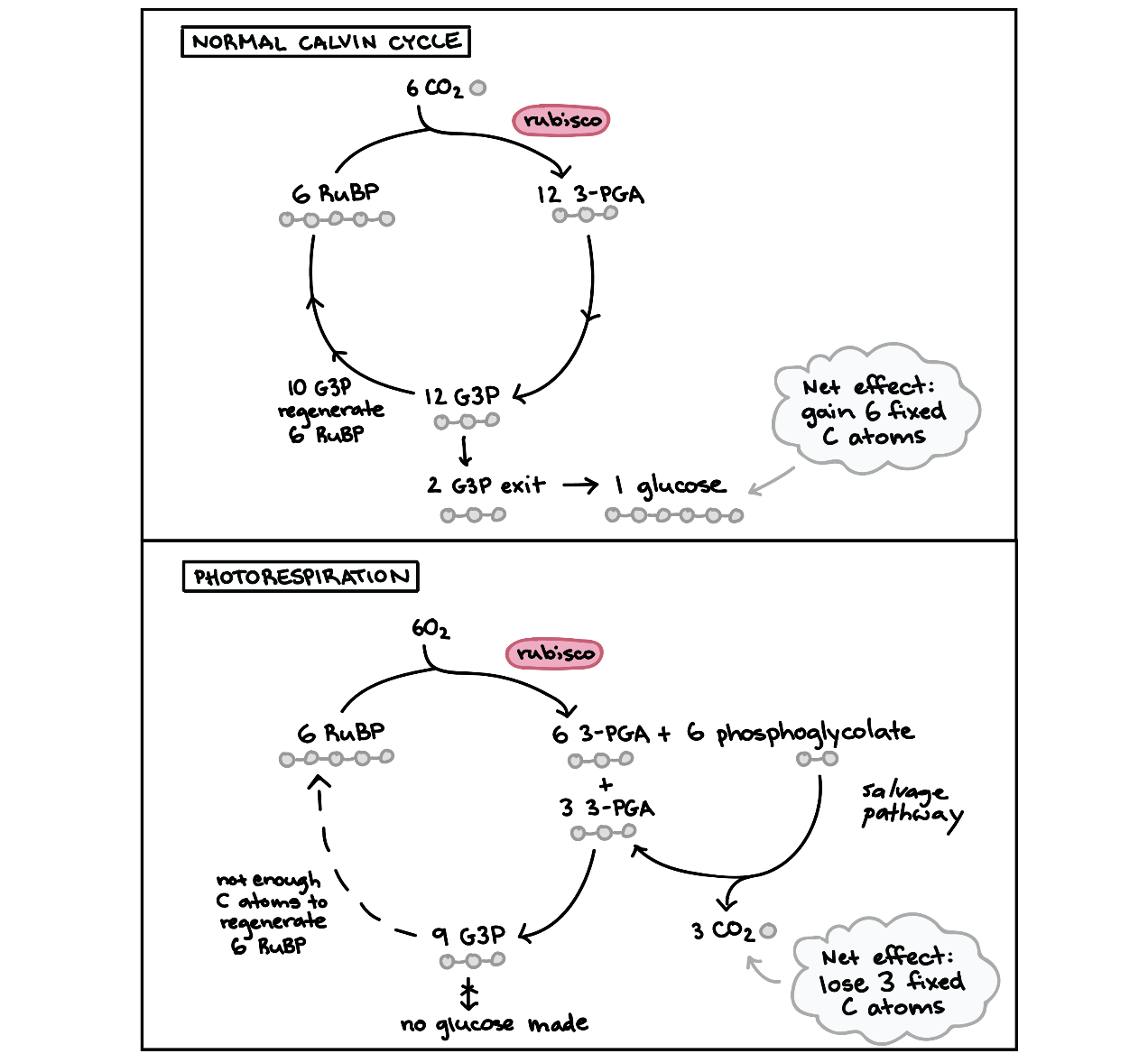
دیدیم که در فتوسنتز از نوع C۳، روبیسکو کربن دی اکسید را طی مرحله اول چرخه کالوین به مولکول آلی تبدیل میکند اما روبیسکو یک نقص عمده نیز دارد و آن این است که چنانچه غلظت کربن دی اکسید در محیط به کمتر از ۵۰ppm برسد به جای کربن دی اکسید، از اکسیژن استفاده میکند. این واکنش فرعی مسیری به نام تنفس نوری را آغاز میکند که به جای تثبیت کربن، منجر به از دست دادن کربن تثبیت شده به شکل کربن دی اکسید میشود. تنفس نوری انرژی را هدرداده و موجب کاهش تولید قند میشود.
عملکرد روبیسکو
روبیسکو هر کدام از مولکولهای کربن دی اکسید یا اکسیژن را به یک ترکیب ۵ کربنی به نام ریبولوز بیفسفات (PBuR) متصل میکند، واکنشی که از کربن دی اکسید استفاده میکند، اولین مرحله از چرخه کالوین است و به تولید قند منجر میشود اما در واکنشی که از اکسیژن استفاده میشود، اولین مرحله از تنفس نوری است که باعث اتلاف انرژی و اختلاف در چرخه کالوین میشود.
تنفس نوری در کلروپلاست آغاز میشود، هنگامی که در واکنش اکسیژنازی، روبیسکو اکسیژن را به ریبولوز بیفسفات وصل میکند، دو مولکول تولید میشود: ترکیب سه کربنه ۳-PGA و ترکیب دو کربنه فسفو گلیکولات.
مولکول سه کربنه به مرحله میانه چرخه کالوین وارد شده، اما مولکول دو کربنه نمیتواند به چرخه وارد شود، بنابراین دو کربن از چرخه خارج میشود.
برای بازسازی ۲ کربن از دست رفته، گیاهان فسفو گلیکلولات را از طریق مجموعه واکنشهایی بین اندامکهای مختلف انتقال میدهند. ۳/۴ فسفو گلیکولاتها بازسازی شده اما ۱/۴ به شکل کربن دی اکسید از دست داده میشود. در شکل زیر میتوانید مقایسه بین تنفس نوری و چرخه کالوین نرمال را مشاهده کنید.
طبق شواهد، تنفس نوری در تثبیت کربن اختلال ایجاد میکند اما ممکن است فواید دیگری برای گیاهان داشته باشد، شواهدی وجود دارند که نشان میدهد تنفس نوری می تواند از آسیب ناشی از برخورد نور به مولکول های درگیر در فتوسنتز جلوگیری کند؛ همچنین تنفس نوری در حفظ تعادل اکسایش اکسیداسیون در سلول ها و ایمنی گیاه اثرات مثبتی دارد.
وقتی روزنهها و منافذ برگ باز هستند کربن دی اکسید به درون برگ وارد شده و اکسیژن و بخار آب از آن خارج میشود و تنفس نوری در حداقل مقدار خود قرار دارد اما هنگامی که روزنههای گیاه بسته است (مثلاً برای کاهش تعرق) اکسیژن حاصل از فتوسنتز درون برگ راهی برای خروج نداشته و در درون برگ انباشته میشود، در این حالت به دلیل بیشتر شدن نسبت غلظت اکسیژن به کربن دی اکسید تنفس نوری افزایش مییابد.
همچنین در دمای معتدل، تمایل روبیسکو به واکنش با کربن دی اکسید حدوداً ۸۰ برابر بیشتر از اکسیژن است؛ اما هرچه دما افزایش مییابد، تمایل روبیسکو به واکنش با اکسیژن بیشتر میشود. بنابراین همان طور که انتظار داریم، در مناطق گرم و خشک احتمال وقوع تنفس نوری بسیار زیاد است. یکی از سازوکارها برای ممانعت از تنفس نوری در گیاهانی وجود دارد که که گیاهان C۴ معروف اند.
گیاهان C۴
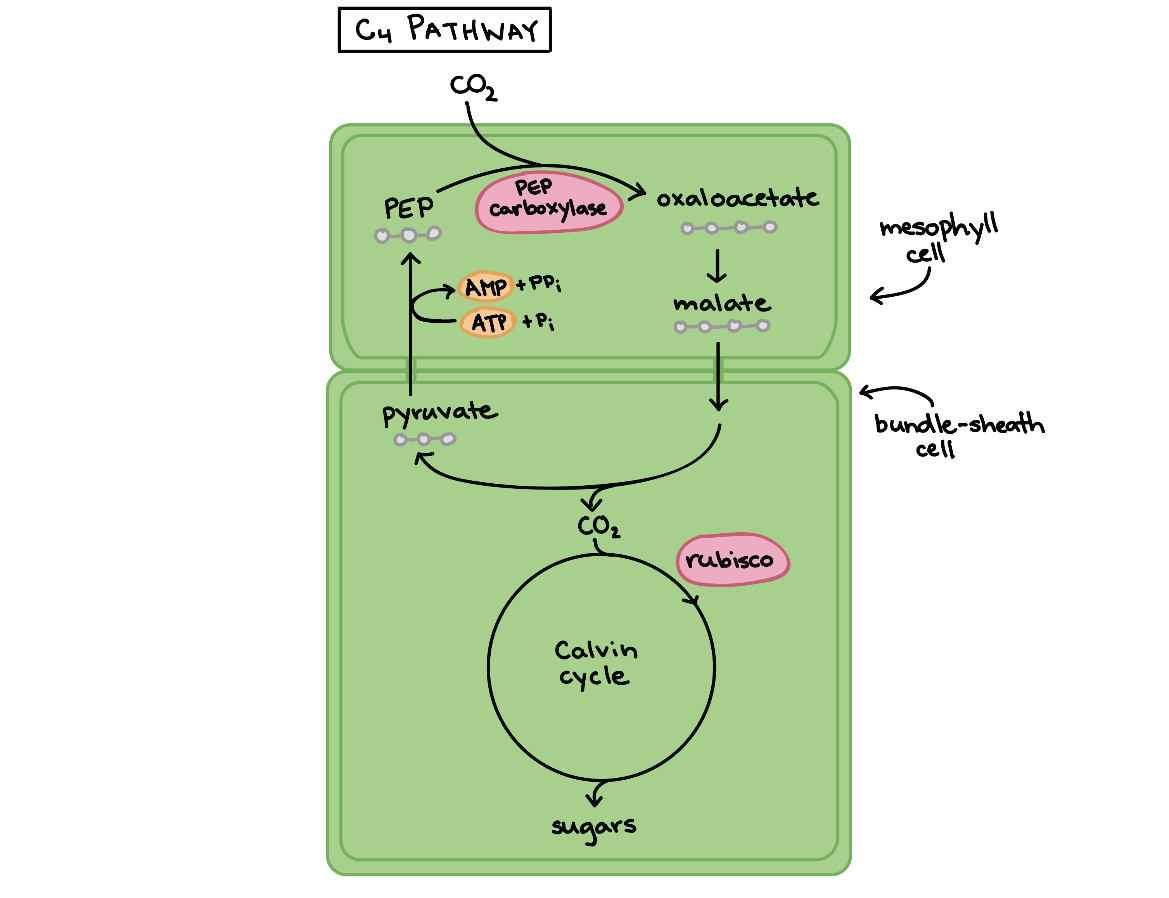
در گیاهان C۴ واکنشهای وابسته به نور و چرخه کالوین از نظر مکانی از هم جدا هستند. واکنشهای نوری در سلولهای پارانشیمی میانبرگ انجام میشود که از دو نوع سلول نردهای و اسفنجی تشکیل شدهاند اما در چرخه کالوین در سلولهای غلاف آوندی انجام میشود.
این تقسیمبندی مکانی به این گونه انجام میشود که ابتدا کربن دی اکسید جو در سلولهای پارانشیمی میانبرگ به شکل یک اسید ساده چهار کربنه (اگزالات) تثبیت میشود. این مرحله توسط آنزیمی غیر روبیسکو به نام PEP کربوکسیلاز انجام میشود که تمایلی به اتصال به اکسیژن ندارد. اگزالات سپس به به مولکول مشابهی به نام مالات که میتواند به سلولهای غلاف آوندی منتقل شود، تبدیل میشود. در درون غلاف آوندی، مالات شکسته شده و از آن مولکول کربن دی اکسید آزاد میشود. کربن دی اکسید آزاد شده توسط روبیسکو تثبیت شده و در چرخه کالوین از آن قند ساخته میشود (همانند فتوسنتز C۳).
این روند بدون صرف انرژی نیست، برای برگرداندن مولکول ۳ کربنی از غلاف آوندی به سلول پارانشیمی و حمل کردن یک کربن دی اکسید دیگر از جو ATP مصرف میشود. سلولهای پارانشیمی، همواره در حال ارسال کربن دی اکسید به سلولهای غلاف آوندی مجاور به شکل مالات است به همین دلیل همیشه غلظت کربن دی اکسید بیشتری نسبت به اکسیژن در اطراف آنزیم وجود دارد که تنفس نوری را به حداقل مقدار ممکن میرساند.
فتوسنتز از نوع C۴ حدودا در ۳٪ گیاهان آوندی مانند ذرت و نیشکر دیده میشود. گیاهان C۴ در مناطق گرم رایجتر است، در محیطهای گرم فواید ناشی از کاهش تنفس نوری از هزینههای صرف شده برای ساخت ATP در گیاه بیشتر است، لذا به نفع گیاهان است که تنفس نوری در آنها به حداقل مقدار ممکن برسد.
از سازوکارهای دیگری که برای جلوگیری از تنفس نوری انجام میشود، در گیاهان CAM است.
گیاهان CAM
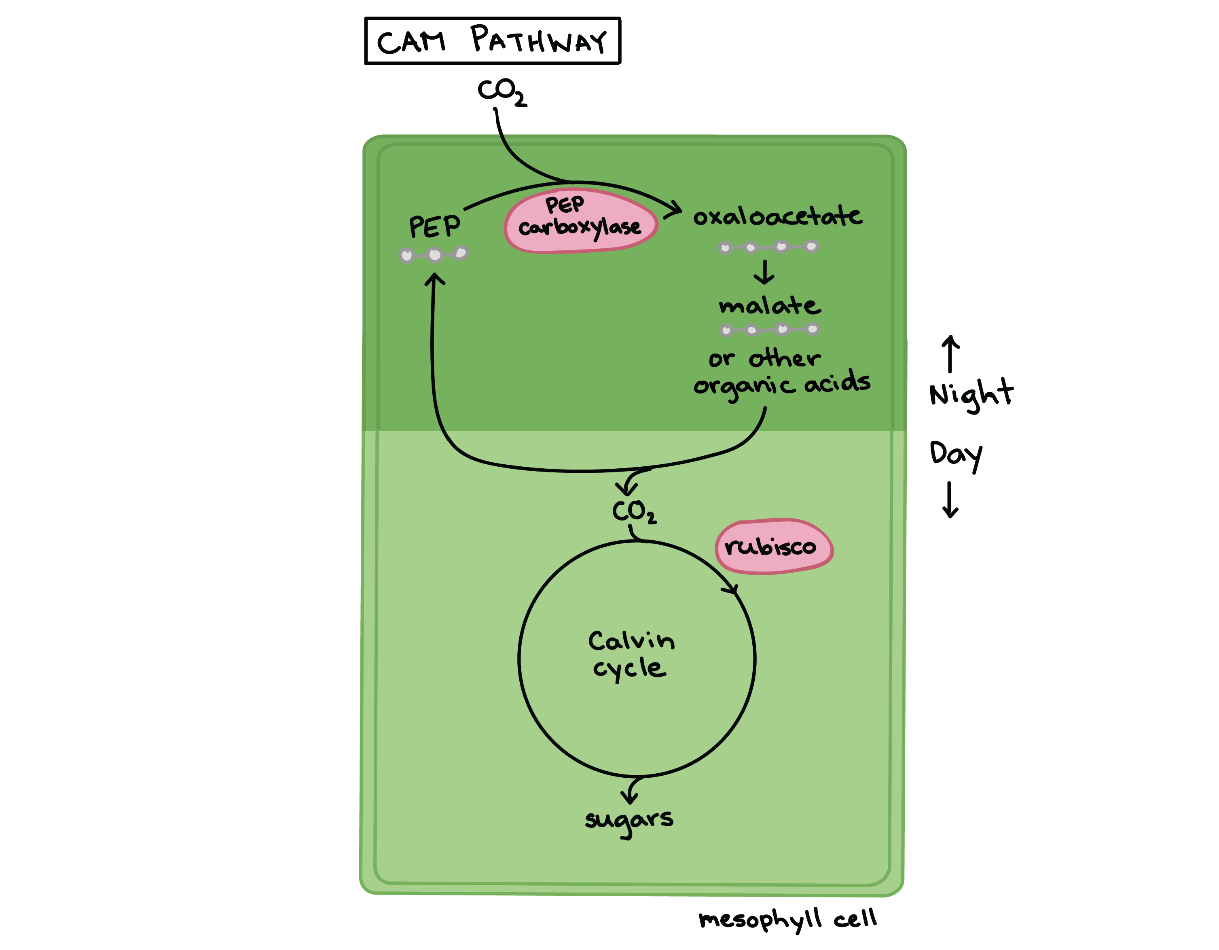
بعضی گیاهان مانند کاکتوس و آناناس در محیطهای خشک سازگارند؛ این گیاهان از متابولیسمهای اسید کراسولاسه استفاده میکنند تا تنفس نوری را به حداقل برسانند. در گیاهان CAM به جای تفکیک مکانی چرخه کالوین و واکنشهای نوری، بین آنها تفکیک زمانی رخ داده است. به این صورت که در شبها گیاهان CAM با باز کردن روزنههایشان به کربن دی اکسید اجازه میدهند که به درون برگها وارد شود. همانند آنچه که در گیاهان C۴ رخ میدهد، این کربن دی اکسید توسط آنزیم PEP کربوکسیلاز تثبیت شده و به اگزالات تبدیل میشود. سپس این اگزالات به مالات یا اسیدهای آلی دیگری تبدیل میشود که آنها تا روز بعد در واکوئلها ذخیره میشوند.
میشوند گیاهان CAM در طول روز، روزنههایشان را باز نمیکنند اما هنوز هم میتوانند فتوسنتز کنند زیرا اسیدهای آلی از درون واکوئل خارج شده و تجزیه میشوند تا کربن دی اکسید مورد نیاز برای چرخه کالوین آزاد شود. این انتشار کربن دی اکسید کنترل شده، غلظت بالایی از کربن دی اکسید را در اطراف روبیسکو حفظ میکند تا تنفس نوری به حداقل مقدار خود برسد. فتوسنتز از نوع CAM همانند گیاهان C۴ در مراحل مختلفی به ATP نیاز دارد. با این حال گونههای گیاهی که از فتوسنتز CAM استفاده میکنند نه تنها از تنفس نوری جلوگیری میکنند بلکه در مصرف آب نیز بسیار کم مصرف هستند!
همانطور که گفته شد روزنه این گیاهان فقط در شب باز میشود؛ هنگام شب هوا خنکتر و رطوبت هوا بیشتر است، این دو عامل، از دست دادن آب از برگها را کاهش میدهد. گیاهان CAM در مناطق بسیار گرم و خشک مانند بیابانها سازگاراند.
مقایسه C۳، C۴، CAM
همه گیاهان C۳، C۴ و CAM از چرخه کالوین برای ساختن قند از مولکول کربن دی اکسید استفاده میکنند روشهای مختلف تثبیت کربن مزایا و معایب مختلفی دارند و گیاهان را برای زیستگاههای مختلف مناسب میسازند. گیاهان C۳ در محیطهای خنک و گیاهان C۴ و CAM در محیطهای گرم و خشک بهترین عملکرد را دارند.
تنفس سلولی، عکس عمل فتوسنتز
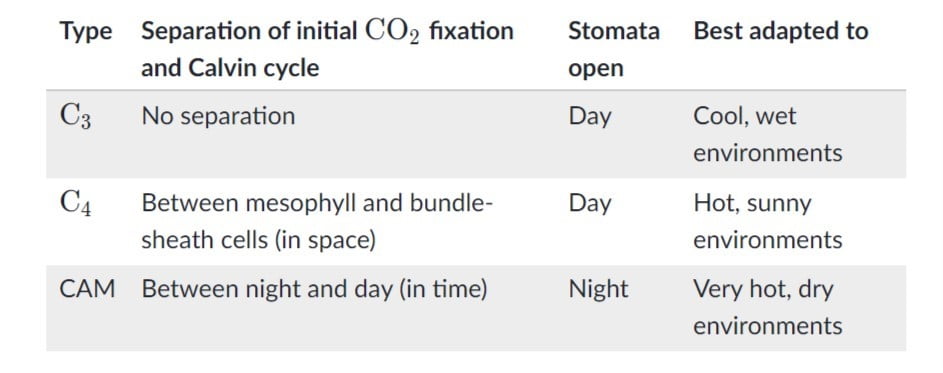
در طی تنفس سلولی یک مولکول گلوکز به تدریج به کربن دی اکسید و آب تجزیه میشود؛ در این فرایند مقداری ATP به طور مستقیم در سری واکنشهای گلیکولیز به وجود میآید اما بیشتر ATP حاصل از تنفس سلولی در فرایند حرکت الکترونها از طریق زنجیره انتقال الکترون در غشای داخلی میتوکندری ساخته میشود.
تنفس سلولی دارای ۴ مرحله است:
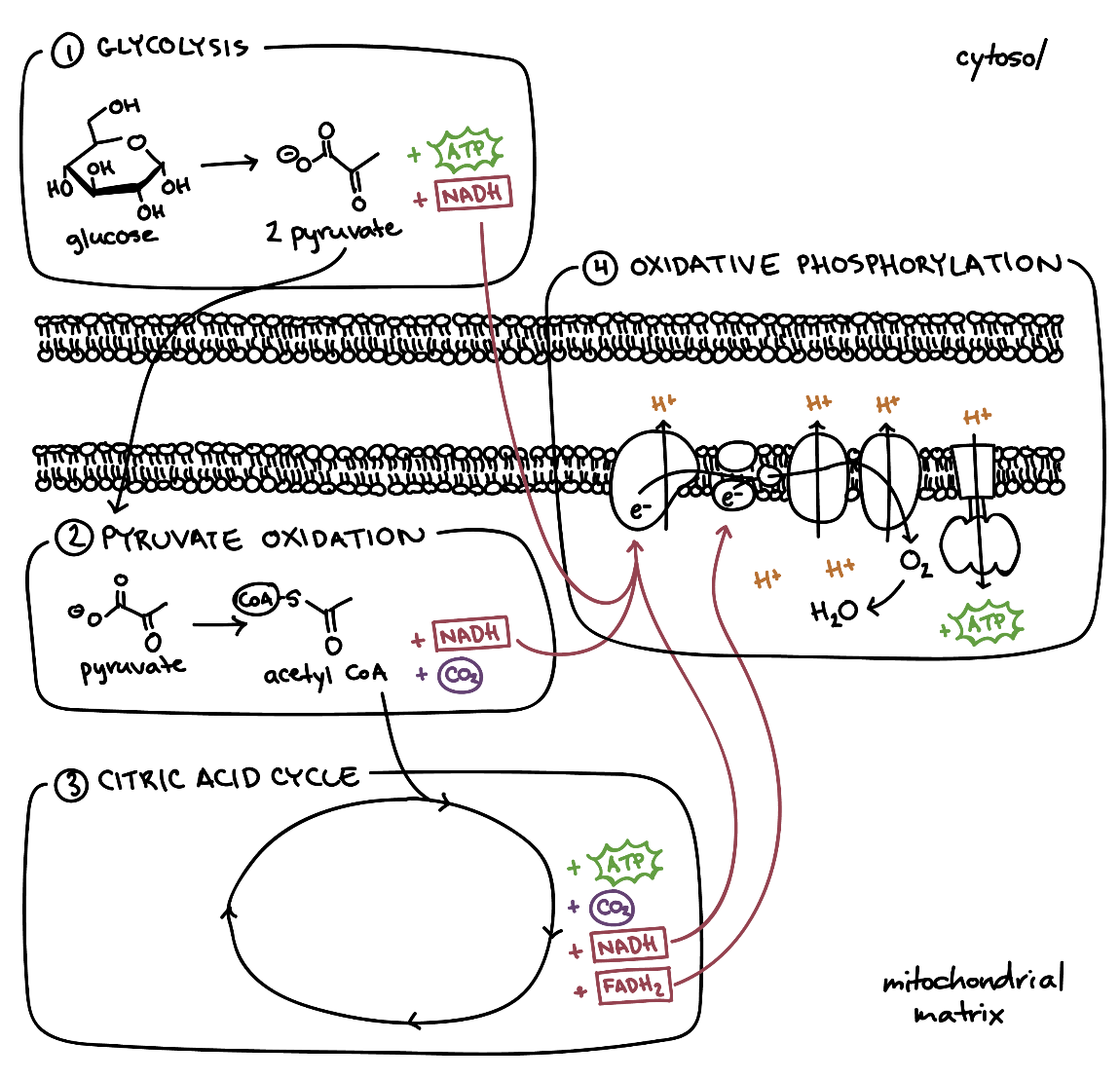
- گلیکولیز: گلیکولیز مجموعه واکنشهایی بیهوازی است که درسیتوپلاسم انجام میشود که طی آن گلوکز به پیروات و یون هیدروژن تبدیل میشود. انرژی آزاد شده در این فرآیند برای تشکیل مولکولهای پر انرژی ATP و NADH استفاده میشود.
- اکسیداسیون پیروات: هر پیروات حاصل از گلیکولیز به ماتریس میتوکندری (داخلیترین فضای میتوکندری) میرود و در آنجا به کوآنزیم A متصل شده و استیل کوآنزیم COA) A) را به وجود میآورد؛ در طی این فرآیند کربن دی اکسید و NADH تولید میشود.
- چرخه کربس: استیل کوآنزیم ساخته شده، با مولکولی چهار کربنه ترکیب شده و در طی چرخه کربس ATP، NADH و FADH۲ را تولید کرده و کربن دی اکسید آزاد میشود. در انتهای این چرخه مولکول ۴ کربنی بازسازی میشود.
- زنجیره انتقال الکترون: در این مرحله NADH و FADH۲ الکترونهای خود را در اختیار زنجیره انتقال الکترون میگذارند و به شکلهای اولیه خود (+NAD و +FAD) بازمیگردند. با حرکت الکترونها در زنجیره، انرژیای آزاد میشود که از آن برای پمپاژ پروتونها از ماتریس به فضای بین دو غشا استفاده میشود. این پمپاژ منجر به ایجاد یک شیب غلظت شده که در آن پروتونها از طریق آنزیمی به نام ATP سنتتاز به داخل ماتریس جریان مییابند و ATP تولید میشود.
از بین مراحل نام برده، گلیکولیز بیهوازی بوده و میتواند بدون اکسیژن هم انجام شود اما سه مرحله دیگر برای وقوع، به اکسیژن نیاز دارند.
تخمیر
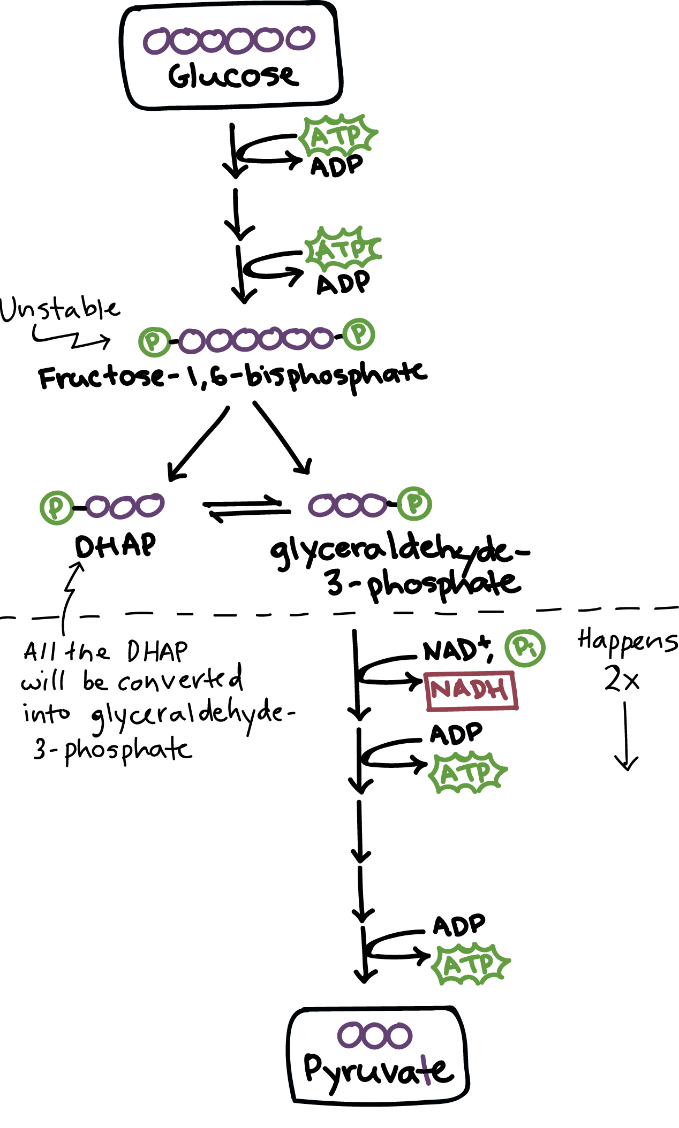
چنانچه اکسیژن در محیط کم باشد یا اصلاً وجود نداشته باشد، انرژی مورد نیاز از طریق فرآیند تخمیر ایجاد میشود. تخمیر فرآیندی متابولیکی است که طی آن مولکولهای آلی (به طور معمول گلوکز) در غیاب اکسیژن یا هر زنجیره انتقال الکترون به اسیدها، گازها یا الکل تبدیل میشوند.
تخمیر، +NAD را که در گلیکولیز برای آزادسازی انرژی به شکل ATP استفاده میشود را، دوباره تولید میکند. با تخمیر فقط دو ATP خالص در هر مولکول گلوکز حاصل میشود (از طریق گلیکولیز)، در حالی که تنفس هوازی با کمک زنجیره انتقال الکترون، ۳۲ مولکول ATP در هر مولکول گلوکز تولید میکند. عملکرد اصلی تخمیر تبدیل NADH به +NAD است تا بتوان از آن برای گلیکولیز دوباره استفاده کرد. در طی تخمیر، یک گیرنده الکترون آلی (مانند پیروات یا استالدهید) با NADH واکنش داده و +NAD را ایجاد میکند و فرآوردههایی مانند دی اکسید کربن و اتانول (تخمیر اتانول) یا لاکتات (تخمیر اسید لاکتیک) تولید میکند.
انواع مختلفی از تخمیر وجود دارد که با محصولات نهایی تشکیل شده از پیروات یا مشتقات آن تمایز می یابند. دو تخمیر که معمولاً مورد استفاده بشر برای تولید محصولات تجاری هستند عبارتند از: تخمیر اتانول (در مشروبات الکلی و نان ) و تخمیر اسید لاکتیک (برای طعم دادن و حفظ لبنیات و سبزیجات).
تخمیر الکلی
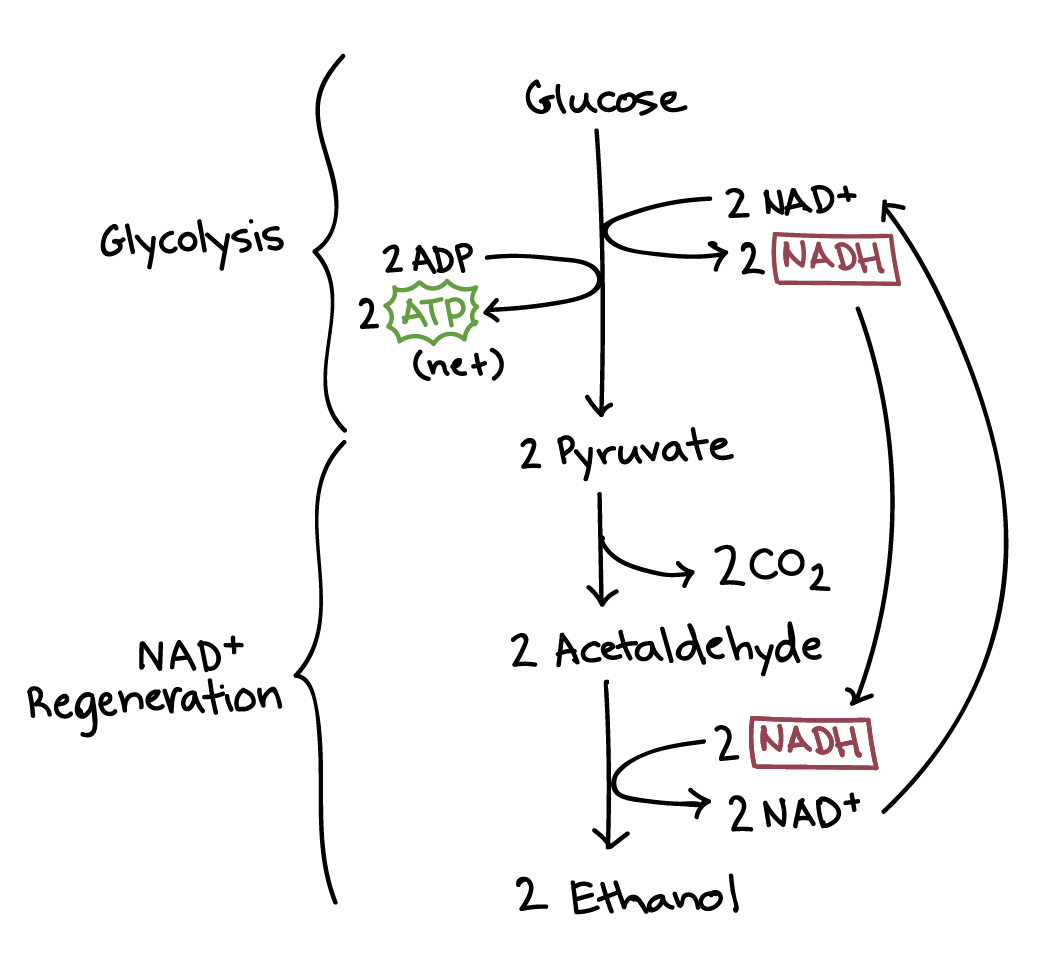
در تخمیر الکلی، پیروات تولید شده از طریق گلیکولیز در دو مرحله به اتانول و دی اکسید کربن تبدیل میشود. ابتدا پیروات با آزادسازی دی اکسید کربن یک ترکیب دو کربنی به نام استالدهید ایجاد میکند. سپس، استالدهید توسط NADH به اتانول کاهش مییابد، در نتیجه +NAD را برای استفاده در گلیکولیز بازسازی میکند. به طور کلی، یک مولکول گلوکز به دو مولکول دی اکسید کربن و دو مولکول اتانول تبدیل میشود.
تخمیر لاکتیکی
دو نوع اصلی از تخمیر لاکتیک اسید وجود دارد: همولاکتیک و هترولاکتیک. در تخمیرهمولاکتیک اسید، NADH پیروات را به طور مستقیم کاهش میدهد و باعث تشکیل لاکتات میشود. در این فرآیند گاز آزاد نمیشود. پس به طور کلی، یک مولکول گلوکز به دو مولکول لاکتات تبدیل میشود. در تخمیر هترولاکتیک، مقداری از لاکتات متابولیزه میشود و در نتیجه اتانول و دی اکسید کربن از طریق مسیر فسفوکتولاز ایجاد میشود. تخمیر اسید لاکتیک در درجه اول توسط انواع خاصی از باکتریها و قارچها انجام میشود. با این حال، هنگامی که اکسیژنرسانی در طی تمرینات شدید کاهش یافته و تنفس هوازی ممکن نباشد، این نوع تخمیر در سلولهای عضلانی نیز برای تولید ATP اتفاق میافتد.

اهمیت ارائه رویکردهای بیوتکنولوژی برای بهبود فتوسنتز
بهبود فتوسنتز با استفاده از رویکردهای بیوتکنولوژی، در تحقیقات بسیاری مورد بررسی قرار گرفته است، ارزیابی این استراتژیها با توجه به شرایط جوی آینده امری حیاتی به شمار میرود. تقاضا برای محصولات زراعی به دلیل افزایش جمعیت، افزایش مصرف کالری و… به طور هشداردهندهای در حال گسترش است.
از گذشته تا کنون بهبود عملکرد فتوسنتز برای دستیابی به محصولات با بازده بالاتر، از اهداف دیرینه متخصصان بیوتکنولوژی گیاهی بوده است؛ با این حال اکثر این مطالعات پیامدهای رشد گیاهان در غلظتهای بالای CO۲ — که در آیندهای نه چندان دور با آن مواجه خواهیم بود — را مد نظر قرار نداده اند. با توجه به اینکه سطح کربن دی اکسید محیط، تاثیر قابل توجهی بر روند فتوسنتز دارد، لذا این ارزیابیها باید با توجه به سناریوهای آینده آب و هوایی بررسی شوند. افزایش ظرفیت فتوسنتز با مهندسی چرخه کالوین در ریز جلبکهای سبز یکی از این سناریوها میباشد. امروزه این تحقیقات با هدف بهبود فتوسنتز و یافتن رویکردهای جایگزینی که موجب افزایش مزایای فتوسنتز شود در حال انجام است.



{kind=link}